Joint work with Simon Levin, Princeton U
Allelopathy in Spatially Distributed Populations
Some strains of E. coli produce a toxic substance, known as
colicins, that kill or inhibit the growth of competing bacteria
of different genotypes, a process known generally as allelopathy .
The strains that produce colicin are immune to its action.
Most theoretical studies of the competition of colicin producing
and colicin sensitive bacteria assume that the population is homogeneously
mixing to arrive at ordinary differential equations
(ODE) describing their dynamics. (See Chao and B.R. Levin (1981)
or Frank (1994).)
If we let u be the density of the colicin producing strain and
v be the density of the colicin sensity strain then we typically have
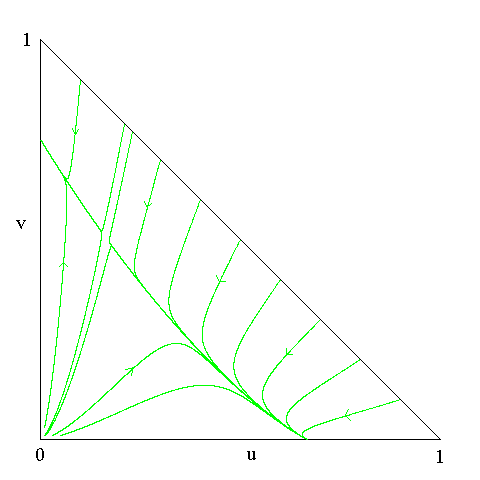
The implication of this picture is that colicin production and
colicin non-production are each evolutionarily stable strategies, i.e.,
a pure population of one type in its equilibrium
cannot be invaded by the other type.
The last conclusion changes if we look at the situation through the
eyes of a spatial model. In our model each point in the square lattice
can be in state 0 = vacant, 1 = occupied by a colicin producer,
2 = occupied by a colicin sensitive bacterium.
Each type is born at empty sites at a rate proportional to the fraction
of the four nearest neighbors occupied by that type. The colicin
producing strain dies at a constant rate 1 while the colicin sensitive
dies at rate 1 plus c times the fraction of colicin producing neighbors.
Because of the last situation it is natural to assume that the birth
rates for the two strains satisfy b1 < b2.
The next four pictures show the evolution of the system with b1=3, b2=4,
c=3 starting from 1% sites of type 1 (red sites) and 50% of type 2
(green sites).
 Time 50
Time 50  Time 300
Time 300
 Time 600
Time 600  Time 900
Time 900
The situation described here is an instance of Case 2 of Durrett and Levin
(1994). The ODE has two attracting fixed
points and predicts that the winner of the competition depends on the
initial densities. In contrast, for the spatial model there is
a stronger type that takes over the system whenever it starts with
a positive density. To be precise we conjecture that the stronger type always
wins if there are infinitely many individuals of that type present
in the initial configuration. Here, by ``wins'' we mean that the probability
a given site x will be occupied by the weaker type converges to 0.
Empirically, the stronger type can be characterized by the behavior of
the system starting from all 2's on half plane x < 0 and all 1's
on the other half. 2's win if the boundary moves to the right
at a linear rate, 1's win if the boundary moves to the left, and
coexistence is never possible.
The last two paragraphs describe the behavior of "Case 2" systems.
Even proving rigorously that the
interface moves at a linear rate with a well defined speed seems to be
an impossibly difficulty problem. However, the idea that interface
speeds identify the stronger equilibrium has been used
by Durrett and Neuhauser (1994) and Durrett and Swindle (1994) to prove
results for particle systems with fast stirring. However, neither that
assumption nor the alternative of long range interactions seems
appropriate in this context.
Chao, L. and B.R. Levin (1981) Structured habitats and the evolution
of anti-competitor toxins in bacteria. Proc. Nat. Acad. Sci.
78 , 6324--6328
Durrett, R. and S.A. Levin (1994) The importance of being discrete
(and spatial). Theoret. Pop. Biol. 46 , 363--394
Durrett, R. and C. Neuhauser (1994) Particle systems and reaction
diffusion equations. Ann. Probab. 22 , 289--333
Durrett, R., and G. Swindle (1994) Coexistence results for catalysts.
Prob. Th. Rel. Fields 98, 489--515
Frank, S.A. (1994) Spatial polymorphism of bacteriocins and other
allelopathic traits. Evolutionary Ecology
This paper is: Durrett, R. and S. Levin (1996)
Allelopathy in Spatially Distributed Populations. Preprint
Back to the Home page