Predator-Prey Models
Part 1: Background: Canadian Lynx and Snowshoe Hares
In the study of the dynamics of a single population, we typically take into consideration such factors as the “natural" growth rate and the "carrying capacity" of the environment. Mathematical ecology requires the study of populations that interact, thereby affecting each other's growth rates. In this module we study a very special case of such an interaction, in which there are exactly two species, one of which -- the predators -- eats the other -- the prey. Such pairs exist throughout nature:
To keep our model simple, we will make some assumptions that would be unrealistic in most of these predator-prey situations. Specifically, we will assume that
Photo source: Rudolfo's Usenet Animal Pictures Gallery (copyright disclaimer).
Very few such "pure" predator-prey interactions have been observed in nature, but there is a classical set of data on a pair of interacting populations that come close: the Canadian lynx and snowshoe hare pelt-trading records of the Hudson Bay Company over almost a century. The following figure (adapted from Odum, Fundamentals of Ecology, Saunders, 1953) shows a plot of that data.
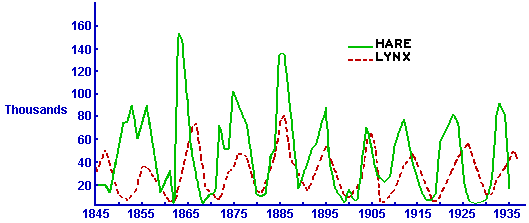
To a first approximation, there was apparently nothing keeping the hare population in check other than predation by lynx, and the lynx depended entirely on hares for food. To be sure, trapping for pelts removed large numbers of both species from the populations -- otherwise we would have no data -- but these numbers were quite small in comparison to the total populations, so trapping was not a significant factor in determining the size of either population. On the other hand, it is reasonable to assume that the success of trapping each species was roughly proportional to the numbers of that species in the wild at any given time. Thus, the Hudson Bay data give us a reasonable picture of predator-prey interaction over an extended period of time. The dominant feature of this picture is the oscillating behavior of both populations.
To be candid, things are never as simple in nature as we would like to assume in our models. In areas of Canada where lynx died out completely, there is evidence that the snowshoe hare population continued to oscillate -- which suggests that lynx were not the only effective predator for hares. However, we will ignore that in our subsequent development.
July 10, 2001
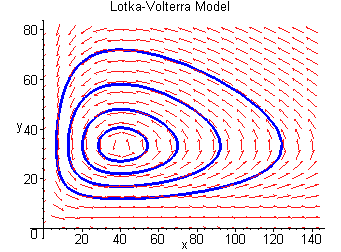
Part 2: The Lotka-Volterra Model
Vito Volterra (1860-1940) was a famous Italian mathematician who retired from a distinguished career in pure mathematics in the early 1920s. His son-in-law, Humberto D'Ancona, was a biologist who studied the populations of various species of fish in the Adriatic Sea. In 1926 D'Ancona completed a statistical study of the numbers of each species sold on the fish markets of three ports: Fiume, Trieste, and Venice. The percentages of predator species (sharks, skates, rays, etc.) in the Fiume catch are shown in the following table:
|
Percentages of predators in the Fiume fish catch |
|||||||||
|
1914 |
1915 |
1916 |
1917 |
1918 |
1919 |
1920 |
1921 |
1922 |
1923 |
|
12 |
21 |
22 |
21 |
36 |
27 |
16 |
16 |
15 |
11 |
As we did with Canadian furs, we may assume that proportions within the "harvested" population reflect those in the total population. D'Ancona observed that the highest percentages of predators occurred during and just after World War I (as we now call it), when fishing was drastically curtailed. He concluded that the predator-prey balance was at its natural state during the war, and that intense fishing before and after the war disturbed this natural balance -- to the detriment of predators. Having no biological or ecological explanation for this phenomenon, D'Ancona asked Volterra if he could come up with a mathematical model that might explain what was going on. In a matter of months, Volterra developed a series of models for interactions of two or more species. The first and simplest of these models is the subject of this module.
Alfred J. Lotka (1880-1949) was an American mathematical biologist (and later actuary) who formulated many of the same models as Volterra, independently and at about the same time. His primary example of a predator-prey system comprised a plant population and an herbivorous animal dependent on that plant for food.
We repeat our (admittedly simplistic) assumptions from Part 1:
If there were no predators, the second assumption would imply that the prey species grows exponentially, i.e., if x = x(t) is the size of the prey population at time t, then we would have dx/dt = ax.
But there are predators, which must account for a negative component in the prey growth rate. Suppose we write y = y(t) for the size of the predator population at time t. Here are the crucial assumptions for completing the model:
![]() These assumptions lead to the conclusion that the negative component of the prey growth rate is proportional to the product xy of the population sizes, i.e.,
These assumptions lead to the conclusion that the negative component of the prey growth rate is proportional to the product xy of the population sizes, i.e.,
Now we consider the predator population. If there were no food supply, the population would die out at a rate proportional to its size, i.e. we would find dy/dt = -cy.
(Keep in mind that the "natural growth rate" is a composite of birth and death rates, both presumably proportional to population size. In the absence of food, there is no energy supply to support the birth rate.) But there is a food supply: the prey. And what's bad for hares is good for lynx. That is, the energy to support growth of the predator population is proportional to deaths of prey, so
![]()
This discussion leads to the Lotka-Volterra Predator-Prey Model:
![]()
![]()
where a, b, c, and p are positive constants.
The Lotka-Volterra model consists of a system of linked differential equations that cannot be separated from each other and that cannot be solved in closed form. Nevertheless, there are a few things we can learn from their symbolic form.
Here is a link for a biological perspective on the Lotka-Volterra model that includes discussion of the four quadrants and the lag of predators behind prey.
Predator-Prey Model, University of Tuebingen, Germany.
Additional links are provided in Part 6 for various extensions of the model.